Краткий обзор проводящих путей спинного и головного мозга

Совокупность аксонов нервных клеток в спинном и головном мозге, которые являются проводниками однородных импульсов, носит название проводящего пути. Все проводящие пути спинного и головного мозга подразделяются на афферентные (восходящие), или центростремительные, эфферентные (нисходящие), или центробежные, и сочетательные, или ассоциативные.
Ассоциативные нервные пути осуществляют связи между нейронами в пределах спинного мозга или той или иной части головного мозга, не выходя за пределы производных каждого мозгового пузыря.
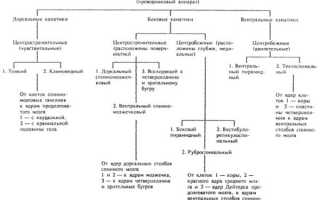
Афферентные, или центростремительные, нервные пути (рис. 947) проводят импульсы от экстеро-, проприо- и интерорецепторов к мозжечку, таламусу, оливам и крыше среднего мозга. Каждый из этих путей проводит импульсы от определенного вида рецепторов.
В спинном мозге восходящие пути образованы аксонами клеток, залегающих в спинномозговых узлах, или аксонами клеток, образующих ядра задних рогов спинного мозга.
Указанные аксоны в составе одних путей проходят в той половине спинного мозга, с которой связаны рецепторы этих путей, в составе других – переходят в другую половину спинного мозга, т. е. происходит перекрест.
В головном мозге восходящие пути состоят из аксонов клеток чувствительных ядер черепных нервов. Эти аксоны по выходе из ядра обычно совершают перекрест – переходят на противоположную сторону.
На пути к мозжечку или ядрам области таламуса центробежные импульсы проходят два нейрона: афферентный, лежащий в периферическом ганглии, и вставочный – в спинном мозге или стволе мозга (продолговатый мозг, мост).
Достигнув области таламуса, восходящие пути заканчиваются на нервных клетках его ядер. В последних локализуются тела третьих нейронов восходящих путей, по которым центростремительные импульсы достигают коры головного мозга.
Участки коры головного мозга, к которым подходят восходящие пути (от органов зрения, вкуса, слуха, обоняния, кожи, внутренних органов), носят название корковых отделов анализаторов: зрительного, вкусового, слухового, обонятельного, кожного, внутреннего (интерорецептивного) и мышечного (двигательного).
Таким образом, в состав анализатора входят афферентные нейроны, нейроны спинного мозга и ствола мозга, а также все клетки коры, участвующие в реакции на импульсы, вызванные раздражением рецепторов.
 Читайте также:
Читайте также:

Эфферентные, или центробежные, пути представляют собой следующее:
- совокупность аксонов определенного вида клеток коры полушарий большого мозга, которые передают нервные импульсы клеткам двигательных ядер черепных нервов или клеткам ядер передних рогов (столбов) спинного мозга;
- совокупность аксонов клеток базальных ядер полушарий большого мозга и ряда ядер ствола, передающих нервные импульсы к эфферентным нейронам спинного мозга и ствола головного мозга.
Первая группа волокон образует пирамидную систему, а вторая – экстрапирамидную систему путей.
Имеются также эфферентные пути вегетативной (автономной) нервной системы, которые из области гипоталамуса передают импульсы на эфферентные нейроны вегетативной (автономной) нервной системы.
Как пути экстрапирамидной системы, так и пути вегетативной (автономной) нервной системы находятся под влиянием коры полушарий большого мозга, которое обеспечивается наличием пучков нервных волокон, соединяющих кору полушарий с базальными ядрами, гипоталамусом и другими ядрами этих систем.
Корковый отдел каждого анализатора состоит из ядра, занимающего определенный участок в коре головного мозга, и рассеянной части – нервных клеток, которые находятся за пределами этих участков.
Ядра двигательного анализатора локализуются в предцентральной извилине, задних отделах средней и нижней лобных извилин. В верхнем отделе предцентральной извилины и в парацентральной дольке находятся корковые отделы двигательных анализаторов мышц нижней конечности, ниже – мышц таза, брюшной стенки, туловища, верхних конечностей, шеи и, наконец, в самом нижнем отделе – головы.
В заднем отделе средней лобной извилины расположен корковый отдел двигательного анализатора сочетанного поворота головы и глаз. Здесь находится и двигательный анализатор письменной речи, имеющий отношение к произвольным движениям, связанным с написанием букв, цифр и других знаков.
Задний отдел нижней лобной извилины является местом расположения ядра двигательного анализатора устной речи.
Корковый отдел обонятельного анализатора (и вкуса) находится в крючке височной доли, зрительного – занимает кору края шпорной борозды (затылочная доля), слухового – в коре средней части верхней височной извилины. В задней части верхней височной извилины расположен слуховой анализатор речевых сигналов (контроль своей и восприятие чужой речи). Зрительный анализатор письменных знаков локализуется в коре угловой извилины.
Корковый отдел анализатора общей чувствительности: температурной, болевой, осязательной, мышечно-суставной – располагается в постцентральной извилине; проекция отдельных частей тела здесь такая же, как и в двигательном анализаторе.
-
Читайте также:

В верхней теменной дольке имеется область коры, обеспечивающая функцию узнавания предметов на ощупь (стереогноз), а в нижней теменной дольке – двигательный анализатор, ответственный за воспроизведение усвоенных в течение жизни координированных движений (праксия, у правшей – слева).
Корковые концы анализаторов связаны с их периферическими отделами (с рецепторами) системой проводящих путей головного и спинного мозга и отходящих от них нервов.
Проводящие пути головного и спинного мозга

В нервной системе импульсы, возникающие при воздействии на рецепторы, передаются по отросткам с нейрона на нейрон. Это осуществляется благодаря синапсам, которые выполняют функцию смыкателя или размыкателя нейронов.
Импульсы передаются только в одном направлении – от рецептора через вставочный нейрон к эфференту, что обусловлено морфофункциональными особенностями синапсов, которые проводят возбуждение только от пресинаптической мембраны к постсинаптической.
Проводящие пути – это совокупность нервных волокон, проходящих в определенных зонах белого вещества головного и спинного мозга, объединенных общностью морфологического строения и функции.
В спинном и головном мозге выделяют по строению и функции три группы проводящих путей.
Ассоциативные пути соединяют участки серого вещества, различные функциональные центры (кора мозга, ядра) в пределах одной половины мозга. Выделяют короткие и длинные ассоциативные волокна. Короткие волокна соединяют близлежащие участки серого вещества и располагаются в пределах одной доли мозга – внутридолевые пучки волокон.
Длинные ассоциативные волокна связывают участки серого вещества, расположенные на значительном расстоянии друг от друга, обычно в различных зонах. К ним относятся верхний продолговатый пучок, соединяющий кору лобной доли с теменной и затылочной, нижний продолговатый пучок, связывающий серое вещество височной доли с затылочной долей.

В спинном мозге ассоциативные волокна связывают между собой нейроны, расположенные в различных сегментах. Они образуют собственные пучки спинного мозга (межсегментарные пучки), которые располагаются вблизи серого вещества.
Короткие пучки перекидываются через 2-3 сегмента, а длинные пучки соединяют далеко расположенные сегменты спинного мозга.
Коммисуральные (спаечные (нервные волокна связывают нервные центры (серое вещество) правого и левого полушария большого мозга, образуют мозолистое тело (коммисуру), спайку свода и переднюю спайку, т.е. коммисуральные волокна проходят из одного полушария в другое. В мозолистом теле располагаются волокна, соединяющие новые, более молодые отделы мозга.
В белом веществе полушарий волокна мозолистого тела расходятся веерообразно, образуя лучистость мозолистого тела.
Проекционные волокна соединяют нижележащие отделы с базальными ядрами и корой, и, наоборот, кору головного мозга, базальные ядра с ядрами мозгового ствола и со спинным мозгом. При помощи проекционных нервных волокон, достигающих кору большого мозга, картины внешнего мира как бы проецируются на кору, как на экран, где происходит высший анализ поступивших импульсов и сознательная их оценка.
Выделяют проекционные восходящие и нисходящие пути. Восходящие (афферентные, чувствительные) несут импульсы, идущие от органов чувств, опорно-двигательного аппарата, внутренних органов и сосудов в головной мозг, к его подкорковым и высшим центрам. По характеру проводимых импульсов восходящие проекционные пути делят на 3 группы:
1) экстерорецептивные пути – импульсы поступают от органов чувств (зрения, слуха, вкуса, обоняния), кожных покровов (болевые, температурные, осязания, давления);
2) проприорецептивные пути – импульсы поступают от органов движения, несут информацию о положении частей тела, о размахе движений;
3) интерорецептивные пути – импульсы поступают от внутренних органов, сосудов (хемо-, баро-, механорецепторы).
Экстерорецептивные проводящие пути. Проводящие пути болевой и температурной чувствительности образуют латеральный (боковой) спинно-таламический путь.
Все восходящие пути состоят из 3 нейронов:
-
Читайте также:

I нейроны располагаются в органах чувств и заканчиваются в спинном мозге или в стволовой части мозга.
II нейроны располагаются в ядрах спинного или головного мозга и заканчиваются в ядрах таламуса, гипоталамуса. Эти нейроны образуют центростремительные восходящие пути.
III нейроны лежат в ядрах промежуточного мозга, для кожной и мышечно-суставной чувствительности – в ядрах таламуса, для зрительных импульсов – в коленчатом теле, для обонятельных импульсов – в сосцевидных телах. Отростки нейронов заканчиваются на клетках соответствующих корковых центров (зрительной, слуховой, обонятельной и общей чувствительности).
Рецепторы первого (чувствительного) нейрона, воспринимающего раздражения, располагаются в коже и на слизистой оболочке, а его тело лежит в спинномозговых узлах; центральный же отросток идет в составе заднего корешка в задний рог спинного мозга.
Аксон второго нейрона, тело которого лежит в заднем роге, направляется на противоположную сторону спинного мозга. Через его переднюю серую спайку аксон входит в боковой канатик, где включается в состав латерального спинно-таламического пути, который поднимается до продолговатого мозга.
Пучок располагается позади оливы, проходит в покрышку моста и покрышку среднего мозга. Аксоны заканчиваются, образуя синапсы на клетках, расположенных в таламусе (III нейрон). Аксоны III нейрона достигают коры полушария, его постцентральной извилины (IV слой коры), где находится корковый конец общей чувствительности.
Импульсы от кожных рецепторов (рецепторы, воспринимающие чувство давления и осязания) поступают к клеткам коры в постцентральную извилину – место общей чувствительности.
Проводящие пути головного и спинного мозга
Проводящие пути — совокупность тесно расположенных нервных волокон, проходящих в определенных зонах белого вещества головного и спинного мозга, объединенных общностью морфологического строения и функции.
Импульсы, возникающие при воздействии на рецепторы, передаются по дендритам нейронов к их телам.
Благодаря многочисленным синапсам нейроны контактируют между собой, образуя цепи, по которым нервные импульсы распространяются только в определенном направлении — от рецепторных нейронов к эффекторным.
Это обусловлено морфофункци-ональными особенностями синапсов, которые проводят возбуждение только от пресинаптической мембраны к постсинаптической мембране.
По одним цепям нейронов импульс распространяется центростремительно: от места его возникновения в коже, слизистых оболочках, органах движения, сосудах к спинному или головному мозгу. По другим цепям нейронов импульс проводится центробежно: от мозга на периферию, к рабочему органу — мышце или железе.
В процессе эволюции возникли сложные рефлекторные дуги, образованные нейронами, расположенными в вышележащих сегментах спинного мозга, в базальных ядрах головного мозга и в коре большого мозга (надсегментарные аппараты).
Отростки нейронов, направляющиеся из спинного мозга к различным структурам головного мозга, а от них в обратном направлении, к спинному мозгу, образуют пучки, соединяющие между собой различные нервные центры.
Эти пучки и составляют проводящие пути.
В спинном и головном мозге выделяют три группы нервных волокон: ассоциативные, комиссуральные и проекционные.
Ассоциативные нервные волокна (короткие и длинные) соединяют между собой группы нейронов (нервные центры), расположенные в одной половине спинного или головного мозга.
Комиссуральные нервные волокна связывают аналогичные центры (серое вещество) правого и левого полушарий большого мозга, образуя мозолистое тело, спайку свода и переднюю спайку. Мозолистое тело соединяет между собой новые, более молодые, отделы коры большого мозга правого и левого полушарий.
Различают восходящие и нисходящие проекционные нервные волокна (проводящие пути). Первые, восходящие, связывают спинной мозг с головным, а также ядра мозгового ствола с базальными ядрами и корой большого мозга. Вторые, нисходящие, идут в обратном направлении.
Проводящие пути спинного и головного мозга

Миелиновые волоконные соединения являются той структурой, которая составляет белое вещество спинного мозга. Они имеют пучковое строение. Каждое такой волоконный компонент бывает:
- укороченным (межсегментарным);
- удлинённым, объединяющего спинной с головным видом мозга и в обратном порядке.
Короткий тип волоконных (ассоциативных) структур обеспечивает связь нервных клеток разных сегментарных зон либо симметричных нервных клеток обратных спинномозговых краёв.
О проводящих путях
Длинный тип волокон разделяются на:
- те, которые восходят, идущие к церебральному органу;
- те, которые нисходят, проходящие от церебральных к спинномозговым образованиям.
Такой волоконный состав формирует проводящие пути спинного мозга.
Аксонный вид пучковых образований формируют около серой мозговой субстанции ряд канатиковых элементов, располагающихся:
- спереди, прилегающие к внутренней стороне от передних роговых структур мозга;
- сзади, локализованных среди задних роговых элементов серого вида мозговой субстанции;
- по бокам, располагающиеся латерально спинномозгового района между корешковыми элементами, находящихся спереди и сзади.
Нервные отростки спинальных узлов и серой субстанции отходят в белую, а потом в разные образования центральной НС.
При этом создаются проводящие пути головного и спинного мозга, которые восходят и нисходят.
О канатиковых образованиях
Нисходящий тип проводящих путей располагается в передних канатиковых элементах:
- пирамидальный тракт, либо передний корковый путь спинного мозга, не являющийся перекрещенным;
- с продольным расположением заднее пучковое образование;
- тектоспинальный тип пути, либо покрышечный тракт спинного мозга;
- преддверный либо вестибулоспинальный.
Задний тип канатиков представлен восходящими путями:
- тонкообразного пучкового образования (пучок Голля);
- пучковая структура клиновидного типа (пучок Бурдаха).
Боковой вид канатиков характерен тем, что там отходят тракты, которые восходят и нисходят.
Пути нисходящего характера состоят из:
- бокового коркового либо пирамидального тракта, являющегося перекрещенным;
- красноядерного либо руброспинального;
- ретикулярного, либо ретикулоспинального.
Проводящие тракты восходящего характера включают в себя:
- спинальный таламический;
- латеральный и спинально-мозжечковые спереди расположенные проводящие спинномозговые пути.
Проприоспинальный (ассоциативный) вид проводящих путей объединяют между собой нервные клетки единичного или различных сегментных спинномозговых зон.
Их начало ─ от нервных волокон серого вида мозгового вещества промежуточного района, проходят в белый вид вещества либо передние канатиковые структуры.
Оканчиваются в сером мозговом веществе промежуточной области либо на моторных нейронах переднероговых иных сегментных зон. Данными связями выполняется ассоциативная функция, заключающаяся в:
- согласовании расположения корпуса тела;
- мышечном тонусе;
- двигательной активности туловищных метамеров.
Проприоспинальные проводящие тракты включают в себя комиссуральный вид волоконных компонентов, который соединяет однородные в функциональном плане несимметрично и симметрично расположенные спинномозговые зоны.
Проводящие тракты нисходящего характера берут своё начало от нервных волокон головного мозга, оканчиваются в спинномозговых нейронных сегментах. К ним относят:
- прямой (передний) и латерально (скрещенный) корковый спинномозговой (от пирамидальных нервных волокон и экстрапирамидальной корковой зоны, которые обеспечивают произвольные движения);
- красноядерный проводящий путь;
- вестибулоспиналный;
- ретикулоспинальный виды путей, регулирующие мышечный тонус.
Объединяет все описанные проводящие тракты то, что конечной их точкой считаются моторные переднероговые нейроны. Пирамидальный проводящий путь заканчивается на моторных нервных волокнах, другого рода тракты заканчиваются в основном на промежуточных нервных клетках.
Латеральный с прямым образуют пирамидный проводящий путь. Латеральный берёт начало от нервных волокон корковой зоны большого мозга. По уровню продолговатого мозгового отдела идёт на противоположный край с формированием перекреста. Затем опускается по другому краю спинного мозга.
Прямая пучковая структура проводящего тракта опускается к своей сегментной области, идёт к моторным нервным волокнам другого края. Из этого следует, что пирамидальная система считается перекрещенной.
Из красных ядер нервных отростков состоит красноядерный спинномозговой тип проводящего пути.
Данные аксонные структуры после того, как вышли из ядерной зоны идут на симметричный край, разделяются на 3 пучковых элемента. Первый проходит к спинному мозгу, второй ─ к мозжечковому району.
Последний идёт к ретикулярной формации мозгового ствола. Нервные волокна, начинающиеся с данного проводящего тракта, управляют тонусом мышц.
Руброретикулярный тип пути с рубромозжечковым осуществляют координационную активность пирамидальных нервных мозжечковых клеток, организующих произвольную двигательную активность.
Вестибулоспинальный проводящий тракт берёт начало от нервных клеток латерально расположенного преддверного ядра, которое лежит в продолговатом мозговом отделе. Оно влияет на то, как активны моторные спинномозговые нейроны, отвечает за:
- состояние мышечного тонуса;
- насколько согласованы движения;
- координацию.
Ретикулоспинальный проводящий путь проходит от ретикулярной формационной мозговой структуры к моторным нейронам, через спинной мозг ретикулярной формацией регулируется мышечный тонус.
Деструкция проводящей системы приведёт к патологии двигательного и чувствительного комплекса пониже повреждённой зоны.
При пересечении пирамидальной системы пониже перерезки вызовет гипертонусное мышечное состояние (спинномозговые мотонейроны освобождены от тормозящего воздействия пирамидальных клеточных структур корковой зоны) и в итоге к спастической парализации.
Когда пересечены тракты, отвечающие за чувствительность, то наблюдается полная утрата мышечной, болевой и других видов сенсетивности пониже перерезанного спинномозгового района.
Восходящие проводящие пути присоединяют спинномозговые и церебральные зоны. Они состоят из:
- путей, отвечающих за проприоцептивный вид сенсетивности;
- таламического тракта;
- спинально-мозжечкового и –ретикулярного.
Также передают данные к церебральному органу об проприо-, интеро-, экстерорецептивных раздражающих воздействиях.
Тонкообразный с клиновидным пучком проприоцептивного пути берут своё начало от рецепторных элементов, отвечающих за глубокую чувствительность мышечных, сухожильных волокон, надкостницы, суставных оболочек. Из узлов, которые собирают данные от каудальных участков тела, тазовой зоны, ног берёт начало тонкое пучковое образование.
Из узлов, выполняющих сбор данных от мышечных волокон грудной клетки, рук начинается клиновидная пучковая структура.
От спинального нервного узла аксонны проходят к задним спинномозговым корешкам, к белой субстанции задних канатиковых структур, подымаются к тонкому клиноподобному ядру продолговатого мозгового отдела.
Тут осуществляется начальный переход на новую нервную клетку, потом путь проходит к латерально располагающимся таламусным ядрам другого большого мозгового полушария. Потом идёт переключение на новую нервную клетку, то есть начинается следующее переключение.
От таламусной зоны путь подымается к нервным клеткам четвёртого слоя соматосенсорного района коры. Волоконные структуры данных трактов выполняют отдачу коллатералей в каждом спинномозговом участке, что обеспечивает коррекцию позиции всего тела. Скорость, при которой проводится возбуждение по волоконным структурам этого пути составляет 65-100 м/с.
Спинальный таламический тракт является основным путём кожного вида сенсетивности. Он берёт своё начало от рецепторов, отвечающих за:
- боль;
- тактильность;
- температурную сенсетивность, включая барорецепторы.
Сигналы от кожных рецепторных образований идут к спинному нервному узлу. Потом через заднюю корешковую структуру к заднероговой спинномозговой зоне (при первом переключении).
Сенсетивный ряд заднероговых нервных клеток отсылает нервные отростки на иной спинномозговой край, осуществляет подъём по латеральному канатиковому образованию к области таламуса.
Скорость, при которой проводится сигнал по ним находится в пределах от 1 до 30 м/с (при втором переключении), затем из этой части к сенсорному району коркового слоя большого мозга. Некоторые волокна кожных рецепторов идут к таламусной зоне по передней канатиковой спинномозговой структуре.
Спинально-мозжечковые виды путей находятся в латеральных канатиковых спинномозговых элементах, имеют неперекрещивающийся передний, спинно-мозжечковый тракт и с двукратным перекрещиванием задний спинально-мозжечковый.
Из этого следует, что все пути спинально-мозжечкового рода берут своё начало с левой стороны тела и оканчиваются в левосторонней мозжечковой доле. Также к правой мозжечковой доле приходят данные со своей части тела.
Эти данные идут от сухожильного рода рецепторных формирований Гольджи, тактильных, а также проприорецепторных, барорецепторных структур. Скорость, при которой проводится сигнал по этим путям, составляет от 110 до 120 м/с.
Проводящие пути спинного и головного мозга

Проводящие пути спинного и головного мозга
Проводящие пути – это совокупность тесно расположенных нервных волокон, соединяющих различные центры головного и спинного мозга, проходящих в определённых зонах их белого вещества и проводящих определённые нервные импульсы.
В спинном и головном мозге выделяют три группы проводящих путей (нервных волокон): ассоциативные, комиссуральные и проекционные.
Проекционные нервные волокна соединяют спинной мозг с головным, ядра мозгового ствола с базальными ядрами и корой полушарий большого мозга (восходящие пути), а также головной мозг со спинным (нисходящие пути).
Нисходящие проводящие пути проводят импульсы от коры полушарий большого мозга и подкорковых центров к ядрам мозгового ствола и двигательным ядрам передних рогов спинного мозга. Эти пути разделяются на две группы: пирамидные и экстрапирамидные. Первые являются главными двигательными путями.
Они несут через соответствующие двигательные ядра головного и спинного мозга импульсы из коры полушарий большого мозга к скелетным мышцам головы, шеи, туловища, конечностей. Экстрапирамидные пути – это рефлекторные двигательные пути.
Они несут импульсы от подкорковых центров и различных отделов коры к двигательным ядрам черепных и спинномозговых нервов, затем к мышцам, а также другим нервным центрам ствола головного мозга и спинному мозгу.
Нисходящие двигательные пути заканчиваются на периферических мононейронах спинного мозга посегментно, они оказывают существенное влияние на его рефлекторную деятельность.
Главный двигательный, или пирамидный корково-спинномозговой путь представляет собой систему нервных волокон, по которым произвольные двигательные импульсы от гигантопирамидальных невроцитов (пирамидных клеток Беца), расположенных в коре предцентральной извилины (5-ый слой) и околоцентральной дольки, направляются к двигательным ядрам черепных нервов и к передним рогам спинного мозга, а от них к скелетным мышцам. В зависимости от направления и расположения волокон пирамидный путь делится на три части: корково-ядерный путь, идущий к ядрам черепных нервов; латеральный и передний корково-спинномозговые пути, идущие к ядрам передних рогов спинного мозга.
Корково-ядерный путь представляет собой пучок аксонов гигантопирамидальных клеток предцентральной извилины. Этот путь начинается в нижней трети предцентральной извилины и проходит через колено внутренней капсулы, основание ножки мозга.
Волокна корково-ядерного пути переходят на противоположную сторону к двигательным ядрам черепных нервов, где заканчиваются синапсами на их нейронах.
Аксоны двигательных нейронов указанных ядер выходят из мозга в составе соответствующих черепных нервов и направляются к скелетным мышцам головы и шеи.
Латеральный и передний корково-спинномозговые (пирамидные) пути начинаются от гигантопирамидальных невроцитов предцентральной извилины.
Волокна этого пути направляются к внутренней капсуле, проходят через переднюю часть её задней ножки, затем через основание ножки мозга и моста, переходят в продолговатый мозг, образуя его пирамиды.
На границе продолговатого и спинного мозга часть волокон корково-спинномозгового пути переходит на противоположную сторону, продолжается в боковой канатик спинного мозга (латеральный корково-спинномозговой путь) и заканчивается в передних рогах спинного мозга синапсами на их двигательных клетках.
Таким образом боковой корково-спинномозговой путь лежит в боковом канатике, состоит из нейритов клеток коры противоположного полушария и постепенно истончается, так как в каждом сегменте спинного мозга часть его волокон заканчивается на клетках передних рогов. Этот путь проводит от коры произвольные двигательные импульсы, стимулирующие и тормозные.
Волокна корково-спинномозгового пути, не переходящие на противоположную сторону на границе продолговатого мозга со спинным, спускаются вниз в составе переднего канатика спинного мозга, образуя передний корково-спинномозговой путь.
Эти волокна посегментно переходят на противоположную сторону через белую спайку спинного мозга и заканчиваются синапсами на двигательных невроцитах (мотонейронах) передних рогов противоположной стороны спинного мозга.
Аксоны клеток передних рогов выходят из спинного мозга в составе переднего корешка и иннервируют скелетные мышцы. Итак, все пирамидные пути являются перекрещёнными.
Представляет интерес, что корково-спинальные пути оканчиваются на мотонейронах спинного мозга лишь у человека и приматов, в то время как у субприматов, а иногда и у приматов между ними включается вставочный нейрон.
При поражении пирамидных путей рефлекторные механизмы спинного мозга растормаживаются, наблюдается усиление рефлексов спинного мозга и тонуса мышц, выявляются защитные рефлексы, а также рефлексы, которые в норме наблюдаются только у грудных детей. Поражение пирмидных путей приводит к развитию центральных параличей или парезов.
Экстрапирамидные пути у млекопитающих и человека являются морфологической основой, по которой проводятся безусловные рефлексы, регулирующие тонус скелетных мышц и осуществляющие их непроизвольную автоматическую иннервацию. При поражении этих путей возникают различного рода гиперкинезы, акинезы.
Экстрапирамидные проводящие пути являются филогенетически более старыми, чем пирамидные. Они имеют множество связей с клетками и ядрами ствола мозга и с корой большого мозга, которая контролирует и управляет экстрапирамидной системой.
В связи с этим общим началом экстрапирамидных путей можно считать кору полушарий большого мозга, а местом, где они оканчиваются, — ядра мозгового ствола и передних рогов спинного мозга.
Влияние коры полушарий большого мозга осуществляется через ряд образований: мозжечок, красные ядра, ретикулярную формацию, связанную с таламусом и полосатым телом через вестибулярные ядра.
Экстрапирамидные пути разделяют на три части: корковые, стриопаллидарные и трункоспинальные пути.
Корковые экстрапирамидные пути слагаются из нервных волокон, идущих от клеток корковых двигательных центров к образованиям экстрапирамидной системы. Здесь можно выделить следующие пути: корково-таламические, корково-гипоталамические, корково-мостовые, корково-красноядерные и корково-покрышечные.
Стриопаллидарные пути представлены нейритами клеток, залегающими в подкорковых базальных ядрах (в полосатом теле – в хвостатом ядре, бледном шаре, скорлупе); эти пути идут к ядрам таламуса, гипоталамуса, красного ядра, чёрного вещества. Анатомически указанные нейриты образуют три основных эфферентных пучка: чечевицеобразная петля, чечевицеобразный пучок и субталамический пучок.
Трункоспинальные пути образуются нервными проводниками, идущими от ядер среднего, промежуточного и продолговатого мозга к двигательным ядрам спинного мозга и черепных нервов, в составе следующих анатомически обособленных двигательных путей.
Одной из функций красного ядра является поддержание мышечного тонуса, необходимого для непроизвольного удержания тела в равновесии. От красного ядра нервные импульсы направляются в двигательные ядра передних рогов спинного мозга (красноядерно-спинальный путь).
Он начинается из среднего мозга (от красного ядра), спускается по боковому канатику противоположной стороны спинного мозга и оканчивается на двигательных нейронах передних рогов.
Этот путь несёт непроизвольные двигательные импульсы, он имеет важное значение для экстрапирамидного обеспечения движений.
Покрышечно-спинномозговой путь начинается от ядер покрышки четверохолмия и заканчивается у клеток передних рогов шейных сегментов, устанавливает связи экстрапирамидной системы, а также подкорковых центров зрения и слуха с шейной мускулатурой; таким образом он связан со слуховыми и зрительными восприятиями.
Оливоспинномозговой путь присутствует в шейных сегментах спинного мозга; начинается от нейронов оливы и заканчивается на клетках передних рогов.
Спинно-ретикулярный путь идёт от ретикулярной формации ствола головного мозга к мотонейронам спинного мозга, осуществляет большое влияние на функции спинного мозга.
Задний продольный пучок начинается от ядра Даркшевича и заканчивается посегментно у мотонейронов спинного мозга. Имеет связи со всеми ядрами глазодвигательных нервов и вестибулярного нерва. Обеспечивает одновременность поворота глазных яблок и головы, содружественность движений глазных яблок. Этот пучок называют также пучком Шютца.
Медиальный продольный пучок лежит в переднем канатике и состоит как из нисходящих, так и восходящих волокон; берёт начало и оканчивается на ядрах ствола мозга и на клетках передних рогов; иннервирует мышцы шеи. Пучок представляет собой очень древнюю систему волокон, которая у низших позвоночных служит важнейшим ассоциационным путём головного мозга.
Все эти проводящие пути функционально объединяют организм в единое целое и обеспечивают согласованность его действий.